Dr Robert Burek,
mgr inż. Dagmara Telus
Ultrasłabe prosięta – jak je uratować?
(Każde prosię na wagę złota)
Postęp genetyczny w produkcji trzody chlewnej w ostatnich dekadach doprowadził do istotnego wzrostu plenności loch. Intensywna selekcja w kierunku zwiększenia liczby owulacji, poprawy przeżywalności zarodków oraz wysokiej wydajności rozrodczej sprawiła, że mioty liczące 18-20 żywo urodzonych prosiąt przestały być wyjątkiem, a w wielu nowoczesnych stadach coraz częściej stają się normą.
Jednocześnie wzrost liczebności miotów niesie za sobą nieuniknione konsekwencje biologiczne. Ograniczona pojemność macicy, konkurencja płodów o składniki odżywcze oraz zmienność warunków rozwoju wewnątrzmacicznego prowadzą do obniżenia średniej masy urodzeniowej prosiąt, zwiększenia ich zróżnicowania i wzrostu odsetka osobników o bardzo niskiej urodzeniowej masie ciała. W praktyce oznacza to większy udział prosiąt słabych, mniej żywotnych i obarczonych wysokim ryzykiem upadku w pierwszych godzinach i dniach życia.
W warunkach licznych miotów klasyczne podejście oparte na „średniej masie prosięcia” traci na znaczeniu. Coraz częściej uwaga hodowców koncentruje się na tzw. prosiętach krytycznych – osobnikach o niskiej masie urodzeniowej, ograniczonych rezerwach energetycznych i obniżonej zdolności do szybkiego pobrania siary. To właśnie przeżywalność w tej grupie decyduje w największym stopniu o wynikach odchowu oraz końcowym efekcie ekonomicznym. Zarządzanie prosiętami krytycznymi jest obecnie jednym z kluczowych wyzwań w nowoczesnej produkcji trzody chlewnej. Nawet niewielka poprawa ich przeżywalności przekłada się bezpośrednio na wzrost liczby prosiąt odsadzonych od lochy oraz obniżenie kosztu produkcji jednego prosięcia.
Tabela 1. Nowa klasyfikacja prosiąt po urodzeniu
| Kategoria prosięcia Masa urodzeniowa Prosięta prawidłowe >1,1 kg Prosięta niskiej masy 0,8-1,1 kg Prosięta ultrasłabe (krytyczne) <800 g |
Pierwsze 6 godzin życia – okno ratunkowe
Badania naukowe, jak również praktyka zootechniczna jednoznacznie wskazują, że pierwsze 6 godzin po porodzie stanowi krytyczne „okno metaboliczno-immunologiczne” decydujące o przeżyciu prosiąt o niskiej masie urodzeniowej oraz prosiąt o niskiej witalności. Pierwsze godziny życia mają decydujący wpływ na to czy prosię:
- uzyska równowagę energetyczną, niezbędną do utrzymania temperatury ciała i aktywności ruchowej,
- będzie w stanie aktywnie dotrzeć do sutka i skutecznie ssać, konkurując z silniejszym rodzeństwem,
- uzyska odporność bierną poprzez pobranie odpowiedniej ilości siary.
Nowe podejście fizjologiczne jasno wskazuje, że energia jest czynnikiem pierwotnym: prosię musi najpierw uzyskać minimalny poziom dostępnej energii metabolicznej, aby być zdolne do skutecznego ssania. Prosię z głębokim deficytem energetycznym, wychłodzone i osłabione nie jest fizjologicznie zdolne do aktywnego pobrania siary, nawet jeśli ma do niej dostęp (Quesnel i wsp. 2021, Devillers i wsp. 2021, Farmer i wsp. 2023).
Filary protokołu ratunkowego „0-24 h”
FILAR 1 – Energia (0-2 h)
Najważniejszym celem noworodka w pierwszych minutach życia jest szybkie pobranie łatwo dostępnej energii. Nowo narodzone prosięta, szczególnie ultrasłabe, rodzą się z minimalnymi zapasami tłuszczu i glikogenu. Bez szybkiego wsparcia energetycznego są słabo zdolne do utrzymania temperatury ciała, aktywnego poruszania się i skutecznego ssania siary.
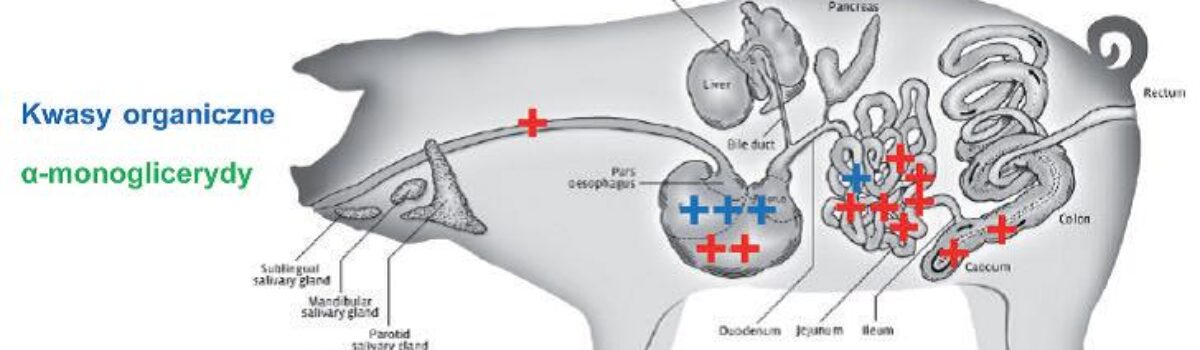
Średniołańcuchowe kwasy tłuszczowe i monoglicerydy tych kwasów – natychmiastowe źródło energii
Średniołańcuchowe kwasy tłuszczowe (MCFA) oraz monoglicerydy tych kwasów stanowią jedno z najszybciej dostępnych źródeł energii dla nowo narodzonych prosiąt, szczególnie tych o niskiej masie urodzeniowej i obniżonej witalności. Ich wyjątkowa wartość wynika z faktu, że są one metabolizowane niezależnie od lipazy trzustkowej, której aktywność u noworodków jest jeszcze niedojrzała. Dzięki temu MCFA mogą być szybko wchłaniane i wykorzystywane energetycznie już w pierwszych minutach życia.
Dodatkową zaletą tych kwasów tłuszczowych jest ich bezpośredni transport do wątroby przez żyłę wrotną, co pozwala na niemal natychmiastowe przekształcenie ich w dostępną energię metaboliczną. Energia ta trafia bezpośrednio do kluczowych narządów, takich jak mózg, serce i mięśnie, umożliwiając szybkie przywrócenie podstawowych funkcji życiowych. W efekcie obserwuje się dynamiczny wzrost dostępnej energii metabolicznej, co sprzyja stabilizacji temperatury ciała, poprawie aktywności ruchowej oraz powrotowi odruchu ssania.
Z praktycznego punktu widzenia podanie MCFA w ciągu pierwszych 30-60 minut po porodzie odgrywa kluczową rolę w ograniczaniu wczesnych strat prosiąt. Tak wczesna interwencja pozwala na szybką stabilizację termiczną noworodka, przywrócenie zdolności do aktywnego ssania oraz znaczące zwiększenie szans na pobranie odpowiedniej ilości siary. W konsekwencji poprawia się skuteczność transferu odporności biernej, a ryzyko upadków w pierwszych godzinach i dniach życia ulega istotnemu ograniczeniu.
FILAR 2 – Odporność (2-12 h)
Po wczesnym ustabilizowaniu statusu energetycznego nowo narodzonego prosięcia możliwe staje się skuteczne wsparcie jego odporności. Kluczową rolę w tym procesie odgrywa siara lochy, która jest jedynym źródłem immunoglobulin – IgG, IgA oraz IgM – niezbędnych do ochrony noworodka przed patogenami środowiskowymi oraz do prawidłowego rozwoju odporności zarówno ogólnoustrojowej, jak i jelitowej. Ponieważ u świń nie dochodzi do przenoszenia przeciwciał przez łożysko w czasie ciąży, całość odporności biernej musi zostać pobrana właśnie w pierwszych godzinach życia.
Z praktycznego punktu widzenia immunoglobuliny powinny być podawane dopiero po wcześniejszym zapewnieniu prosięciu odpowiedniego poziomu energii metabolicznej. Prosię osłabione, wychłodzone i z deficytem energetycznym nie jest fizjologicznie zdolne do aktywnego ssania ani do efektywnego pobrania siary, nawet jeśli jest ona dostępna. Dlatego wsparcie energetyczne stanowi warunek wstępny skutecznego transferu odporności biernej.
Najbardziej pożądanym źródłem przeciwciał pozostaje siara własnej lochy, jednak w praktyce, szczególnie w przypadku bardzo licznych miotów lub prosiąt ultrasłabych, coraz częściej wykorzystuje się także koncentraty serwatkowe oraz proszek jajeczny bogaty w immunoglobuliny jako uzupełnienie naturalnej siary. W ostatnich latach rośnie również znaczenie preparatów łączonych, które dostarczają jednocześnie energii i immunoglobulin. Badania oraz doświadczenia praktyczne wskazują, że takie rozwiązania poprawiają przeżywalność prosiąt w większym stopniu niż podawanie samych immunoglobulin bez wcześniejszej stabilizacji energetycznej.
Liczne prace naukowe potwierdzają, że prosięta ultrasłabe, o masie urodzeniowej poniżej 800 g, które pobierają niewystarczającą ilość siary w pierwszych godzinach życia, charakteryzują się istotnie niższym stężeniem IgG w surowicy oraz gorszymi wynikami przeżywalności. Quesnel i wsp. (2021) oraz Devillers i wsp. (2021) wykazali jednoznacznie, że opóźnione lub ograniczone pobranie siary prowadzi do nieefektywnego transferu odporności biernej i znacząco zwiększa ryzyko upadków w pierwszych dobach życia prosiąt.
FILAR 3 – Jelita i mikrobiota (12-24 h)
Kolonizacja jelit u prosiąt rozpoczyna się już w pierwszej dobie życia i odgrywa kluczową rolę w kształtowaniu ich zdrowia, odporności oraz ryzyka wystąpienia biegunek w kolejnych dniach odchowu. Wczesne zasiedlenie przewodu pokarmowego przez odpowiednie mikroorganizmy wpływa na dojrzewanie bariery jelitowej, funkcjonowanie układu immunologicznego oraz zdolność organizmu do obrony przed patogenami środowiskowymi.
Szczególne znaczenie w tym okresie przypisuje się probiotykom z rodzaju Bacillus, takim jak Bacillus subtilis i Bacillus licheniformis. Bakterie te tworzą przetrwalniki, co istotnie zwiększa ich przeżywalność w niekorzystnych warunkach przewodu pokarmowego noworodków oraz skuteczność działania w jelitach. Dzięki temu mogą one efektywnie kolonizować jelito, wspierać integralność bariery jelitowej oraz ograniczać namnażanie się patogenów, w tym enterotoksycznych szczepów Escherichia coli. Jednocześnie probiotyki typu Bacillus stymulują rozwój odporności miejscowej w obrębie jelit, co ma istotne znaczenie dla ochrony prosiąt w pierwszych dniach życia.
Coraz więcej danych wskazuje również na korzystne, synergistyczne działanie połączenia probiotyków z rodzaju Bacillus ze średniołańcuchowymi kwasami tłuszczowymi (MCFA). Taka kombinacja sprzyja rozwojowi korzystnej mikrobioty jelitowej, stabilizuje funkcje przewodu pokarmowego oraz ogranicza procesy zapalne w obrębie jelit. W praktyce przekłada się to na zmniejszenie ryzyka biegunek oraz redukcję kolonizacji jelit przez patogeny w najbardziej wrażliwym okresie życia prosiąt.
Z punktu widzenia praktyki fermowej oznacza to, że wczesna kolonizacja jelit powinna być traktowana jako jeden z kluczowych elementów protokołu ratunkowego, rozpoczynającego się już w pierwszych godzinach życia. Zastosowanie probiotyków typu Bacillus sprzyja poprawie integralności bariery jelitowej oraz rozwojowi układu odpornościowego, natomiast ich łączenie z MCFA dodatkowo wzmacnia efekt ochronny, ograniczając ryzyko biegunek i negatywnego wpływu patogenów na zdrowie i przeżywalność prosiąt.
Preparat wspierający protokół ratunkowy – Eliksir Życia
Preparat wspierający protokół ratunkowy, taki jak Eliksir Życia oferowany przez Dr Burek Service, może stanowić praktyczną i skuteczną podstawę kompleksowego postępowania z osłabionymi noworodkami. Jego działanie obejmuje jednocześnie wszystkie trzy kluczowe filary wczesnego wsparcia prosiąt. Zawarte w preparacie średniołańcuchowe kwasy tłuszczowe oraz ich monoglicerydy dostarczają natychmiast dostępnej energii, co pozwala szybko ustabilizować funkcje życiowe i przywrócić aktywność osłabionych zwierząt. Obecność immunoglobulin wspiera ochronę przed patogenami w okresie, gdy własna odporność prosięcia jest jeszcze niedojrzała. Z kolei dodatki probiotyczne w postaci Bacillus subtilis i Bacillus licheniformis korzystnie wpływają na kształtowanie mikrobiomu jelitowego, wzmacniając barierę jelitową i ograniczając ryzyko zaburzeń trawiennych.
W praktyce już po podaniu jednej dawki preparatu ultrasłabym prosiętom obserwuje się wyraźną poprawę witalności oraz pojawienie się aktywnego odruchu ssania. Z tego względu Eliksir Życia powinien być stosowany natychmiast po zauważeniu osłabionego prosięcia i stale dostępny w porodówce, aby możliwa była szybka interwencja w krytycznym oknie pierwszych 24 godzin życia.
Wnioski praktyczne
- W stadach o wysokiej plenności najważniejsze jest zarządzanie prosiętami krytycznymi, a nie tylko „średnimi”.
- Pierwsze 6 godzin życia stanowi krytyczne okno energetyczno-immunologiczne – brak szybkiego wsparcia energetycznego prowadzi do hipoglikemii → braku odruchu ssania → braku pobrania siary → wzrostu ryzyka śmierci.
- Kompleksowy protokół ratunkowy powinien uwzględniać dostarczenie energii, stymulowanie wytworzenia odporności biernej i prawidłowego rozwoju przewodu pokarmowego i mikrobioty.
- Stosowanie preparatów będących źródłem natychmiastowej energii, immunoglobulin i probiotyków (np. Eliksir Życia) skutecznie zwiększa przeżywalność ultrasłabych prosiąt i ogranicza wczesne straty w stadzie (fot. 1).
- Istotnym elementem procedury ratowania słabych prosiąt musi być prostota i łatwość aplikacji. Już po podaniu jednej dawki Eliksiru Życia (fot. 2) większość prosiąt nabiera natychmiastowej witalności i chęci do aktywnego ssania.
Naszym celem nie ma być zastąpienie siary, bo nic jej nie zastąpi, a jedynie pomoc prosięciu w chęci jej pobrania. Eliksir Życia jest właśnie takim produktem. Należy jednak pamiętać, że jeżeli prosię nie będzie miało wolnego sutka, to nie napije się siary. Dlatego zarządzanie prosiętami w licznych miotach, których liczebność przekracza liczbę sutków musi uwzględniać „zarządzanie siarą”, to znaczy prosięta, które już opiły się siarą muszą być umieszczone w budkach lub okręgach ograniczających ich dostęp do wymienia. Tylko wtedy kolejne prosięta, które przychodzą na świat mają swój sutek.
Prawdziwym eliksirem jest siara, a Eliksir Życia (Dr Burek Service) jedynie pomaga prosiętom mieć siłę ją pobrać i w konsekwencji wyrosnąć na pełnowartościowego tucznika.
Tabela 2. Filary wsparcia nowo narodzonych prosiąt (0-24 h)
| Filar | Czas interwencji | Cel/funkcja | Środki/metody | Efekt praktyczny | Źródła |
| 1. Energia | 0-2 h | Stabilizacja temperatury, przywrócenie aktywności i odruchu ssania | – MCFA (średniołańcuchowe kwasy tłuszczowe) i ich monoglicerydy | – Szybkie zwiększenie dostępnej energii | Jackman et al. 2020, Philips et al. 2021, Quesnel et al. 2021 |
| – Połączenie MCFA + glicerol | – Poprawa termoregulacji | ||||
| – Przywrócenie odruchu ssania | |||||
| 2. Odporność | 0-12 h | Skuteczny transfer odporności biernej | – Siara lochy | – Wyższe stężenie IgG, IgA, IgM | Quesnel et al. 2021, Devillers et al. 2021, Farmer et al. 2023 |
| – Koncentraty serwatkowe/ proszek jajeczny | – Lepsza odporność ogólnoustrojowa i jelitowa | ||||
| – Preparaty łączone energia + Ig | – Zmniejszenie ryzyka zachorowań i strat | ||||
| 3. Jelita i mikrobiota | 0-24 h | Poprawa integralności bariery jelitowej i stabilizacja mikrobioty | – Bacillus subtilis, Bacillus licheniformis | – Ograniczenie kolonizacji patogenów (E. coli) | Lodemann et al. 2022, Hassing et al. 2023, Zhao et al. 2024 |
| – Połączenie z MCFA | – Zmniejszenie ryzyka biegunek | ||||
| – Wsparcie odporności miejscowej i ogólnej |